Порядок ржавчинные (Uredinales)
Мицелий ржавчинных вполне эндофитный, проходящий по межклетникам, с гаусториями, внедряющимися в полость самих клеток растения.
У большинства форм распространение мицелия ограничено (местный мицелий), у некоторых видов он пронизывает целые побеги взрослого растения (диффузный мицелий). Цитологически мицелий может быть гаплоидным, сложенным из одноядерных клеток, и диплоидным, состоящим преимущественно из двуядерных клеток. В клетках мицелия, особенно в уредоспорах, содержатся капли масла, окрашенные в оранжевый цвет. В связи с этим пятна на растениях, вызванные этими грибами, обычно бывают ржавого цвета, а вызываемые ими болезни названы ржавчиной.
Базидии образуются при прорастании покоящихся спор (телейтоспор), которые, в свою очередь, развиваются на конечных раз-ветвлениях вегетативного мицелия без образования плодовых тел. Базидиоспоры формируются на боковых стеригмах, отходящих по одной от каждой клетки четырехлетней — базидии — фрагмобазидии (эпибазидии).
В цикле развития большинства ржавчинных грибов отмечается правильная и последовательная смена различных спороношений, развивающихся обычно на вегетирующих органах растений (листья, стебли и т. д.). Различают пять таких спороношений и спор, которые часто обозначают цифрами: 0 — спермогонии, или пикниды со спермациями, или пикноспорами; I — эцидии с эцидиоспорами; II —уредопустулы и уредоспоры; III —телейтопустулы и телейтоспоры; IV — базидии и базидиоспоры. При этом спермогонии и эцидии развиваются на гаплоидном мицелии, уредо — и телейтопустулы — на диплоидном, фрагмобазидии с базидиоспорами — на прорастающих телейтоспорах. Все спороношения закладываются эндофитно: в глубине ткани, под эпидермисом, между эпидермисом и кутикулой. Содержащиеся в них споры освобождаются через разрыв тканей растений.
Спермогонии, или пикниды,— шаровидные или кувшиновидные (бывают и плоские) образования, в которых развиваются одноклеточные спермации, или пикноспоры, выходящие с каплями сахаристой и пахнущей жидкости. Они не вызывают дальнейшего заражения (обозначаются нулем), но участвуют в половом процессе ржавчинных грибов. Развиваются они обычно вместе с эцидиями.
Эцидии (Aecidium) — более крупные вместилища чашевидной формы, наполненные золотисто-желтой или оранжевой порошащей массой спор — эцидиоспор, образующихся всегда цепочками. Закладываются эцидии под эпидермисом как шаровидные образования. Затем по мере созревания эцидиоспор эцидии прорываются на поверхность в виде урночки или чаши, наполненной массой эцидиоспор. Самым существенным является то, что у большинства ржавчинных грибов эцидиоспоры окружены особой оболочкой — перидием, состоящим из одного слоя клеток с сильно утолщенными стенками. Эцидии представлены главным образом в семействе пукциниевых (Pucciniaceae), а также мелампсоровых (Меlampsoraceae), паразитирующих на хвойных породах. Эцидиоспоры распространяются воздушными течениями и прорастают всегда в диплоидный мицелий, который проникает в ткани растения через устьица. Кроме собственно эцидия, или эцидиума, у ржавчинных грибов различают еще несколько форм эцидиальных спороношений.
Цеома (Саеота) закладывается под эпидермисом как плоское сплетение, нарастающее краями. Перидий в виде общей оболочки отсутствует, а хорошо развиты парафизы, окружающие эцидиоспоры, которые при созревании разрывают эпидермис и освобождают эпидиоспоры в виде пыльцы оранжево-желтого цвета.
Рестелий (Roestelia) по строению и развитию сходен с эцидием, но отличается сильно развитым перидием, который высовывается из ткани растения в виде рожка или волоска. Внутренняя их полость заполнена эцидиоспорами. Перидий разрывается отверстием с зубцами на вершине, трещинами или щелями. Представлен в роде Gymnosporangium.
Перидермий (Peridermium) имеет очень развитый перидий, который выступает из пораженной ткани в виде пузыревидного или мешковидного вздутия и долго сохраняется. Этот тип эцидиального спороношения представлен у Cronatrium, Coleosporium Melampsoridium.
Они всегда одноклеточные, двуядерные с ростковыми порами, прорастают без периода покоя вегетативно, повторяясь летом несколько раз, гифы проникают в растение через устьице. Уредоспоры называются также летними повторяющимися спорами и спорами размножения, так как они обеспечивают массовое распространение и эпифитотию ржавчины в летний период.
Телейтопустулы, или телейтокучки, образуются на том же мицелии, такой же формы, как уредопустулы, заменяя их к концу вегетационного периода. Телейтоспоры имеют толстую почти черную оболочку и предназначены для сохранения гриба в зимний период, поэтому их называют зимними спорами. Форма и строение телейтоспор у различных представителей очень разнообразны, — эти признаки служат основой для систематики ржавчинных грибов.
У одних видов телейтоспоры одноклеточные, у других двуклеточные, трехклеточные и многоклеточные. Они могут быть на ножках и не склеены между собой, свободные, хотя и располагаются тесным слоем. У многих видов телейтоспоры лишены ножек и соединены в однослойные корочки под эпидермисом или срастаются боками, образуя общую колонку.
Телейтоспоры имеются у всех ржавчинных грибов. Они являются заключительной формой цикла развития, так как в них заканчивается половой процесс, начавшийся в эцидиях и эцидиоспорах. Прорастание телейтоспор у большинства ржавчинников осуществляется после зимовки фруктификативно, т. е. с образованием базидий и базидиоспор. У разных видов телейтоспоры прорастают по-разному.
Базидии образуются при прорастании телейтоспор, каждая клетка которой дает одну фрагмобазидию с четырьмя базидиоспорами. Базидиоспоры одноклеточные, гаплоидные, после созревания отбрасываются от базидии, разносятся ветром, попадают на соответствующие растения и прорастают вегетативно с образованием мицелия. В отличие от других спороношений базидиоспоры дают мицелий, непосредственно проникающий в ткань растения, прорывая кутикулу. Они вызывают первое весеннее заражение растений.
Перечисленные спороношения, в правильной последовательности следующие друг за другом, не полностью представлены у всех ржавчинных грибов. Одни из них, имеющие все указанные спороношения в цикле развития, называются полными; другие ржавчинные грибы, у которых выпали какие-либо спороношения, называются неполными, или сокращенными. В свою очередь эти спороношения могут развиваться на одном или на различных растениях. Ржавчинные грибы, у которых весь цикл развития (гаплоидный и диплоидный мицелий и все спороношения) проходит на одном растении, называются однохозяйными. У других ржавчинных грибов часть цикла (гаплоидный мицелий с эцидиями и спермогониями) развивается на одном виде растения, а вторая часть (диплоидный мицелий с уредо — и телейтопустулами) — на другом виде растения, часто систематически далеком от первого. Такие грибы называются разнохозяйными.
Ржавчинные грибы, как облигатные паразиты, отличаются строгой специализацией, вследствие чего ботанический вид распадается на ряд специальных форм (forma specialis), приуроченных к паразитизму на определенных видах растений. Эти специальные формы состоят из большого количества еще более мелких единиц — биотипов, способных поражать только определенные сорта или группы растений.
Ржавчинные грибы составляют наиболее крупный порядок класса базидиомицетов. На основе строения телейтоспор эти грибы делят на два семейства: пукциниевые (Pucciniaceae) — телейтоспоры на ножках; мелямпсоровые (Melampsoraceae) — телейтоспоры без ножек.
В семействе пукциниевых (Pucciniaceae) телейтоспоры одноклеточные, двуклеточные и многоклеточные, различной формы, образующие рыхлые, пылящие или плотные подушечки (пустулы). Эцидии с перидием или без перидия.
Однохозяйные или разнохозяйные формы отмечаются преимущественно на покрытосеменных. Многие имеют большое практическое значение как возбудители болезней зерновых, бобовых, технических культур.
Uromyces pisi Schroet — возбудитель ржавчины, гороха, полный разнохозяйный паразит. Телейтоспоры одноклеточные. Эцидии и спермогонии развиваются на видах молочая, который под воздействием гриба сильно деформируется, уредо — и телейтоспоро- ношения — на горохе и других бобовых.
Uromyces betae Lev.— полный однохозяйный вид, возбудитель ржавчины свеклы.
Puccinia graminis Pers.— возбудитель стеблевой ржавчины злаков. Телейтоспоры двуклеточные. Полный разнохозяйный вид.
Эцидии и спермогонии на барбарисе, уредо — и телейтоспороношения на злаках. Возбудитель стеблевой ржавчины является сложным видом, в состав которого входят несколько специализированных форм.
Puccinia coronifera Kleb.— возбудитель корончатой ржавчины овса. Эцидии и спермогонии на слабительной крушине (Rhamnus cathartica), уредо — и телейтостадии — на овсе.
Puccinia coronata Corda — возбудитель корончатой ржавчины злаков. Эцидии развиваются на крушине ломкой (Frangula alnus), уредо — и телейтостадии на различных злаках, в том числе растущих в лесах.
Puccinia ribesii—caricis Kleb — возбудитель бокальчатой ржавчины смородины и крыжовника. Полный разнохозяйный паразит. Эцидии развиваются на смородине, крыжовнике и других видах Ribes, уредо — и телейтостадии — на видах осок.
Tranzchelia prunispinosae Dietr.— полный разнохозяйный паразит. Эцидии и спермогонии развивает на Anemone ranunculoides на диффузном мицелии, зимующем в корневищах, уредо — и телейтостадии — на листьях терна (Prunus spinosa). Уредоспоры перемешаны с парафизами. Телейтоспоры двуклеточные, на ножках, сильно перетянуты на уровне перегородки, с бурой бородавчато-шиповатой оболочкой, без пор. Близкий вид Т. fussa Wint. не имеет эцидия, это микроформа.
Gymnosporangium sabinae Wint. Телейтоспоры двуклеточные, с длинной ножкой, погруженной в общую слизистую оболочку. Эцидий типа рестелия, конический, перидий которого при созревании разрывается продольными полосами. Неполный разнохозяйный ржавчинный гриб — отсутствует уредостадия. Эцидии развиваются на гипертрофированных пятнах листьев груши, телейтостадия — на стеблях казацкого можжевельника (Juniperus sabinae), где вызывает значительное утолщение (гипертрофию).
Gymnosporangium Juniperi Link. Эцидии на листьях рябины (Sorbus aucuparia), где вызывает гипертрофированные пятна. Телейтоспоры на молодых ветках обыкновенного можжевельника (Juniperus communis), где вызывает плоские подушечки спороношения.
Gymnosporangium juniperinum Martius (G. tremalloides Hartig). Эцидии на листьях яблони, перидий разрывается на вершине. Их появление приурочено к концу лета и мало вредоносно. Телейтоспоры на обыкновенном можжевельнике (Juniperus communis), вызывают гипертрофию молодых ветвей.
Gymnosporangium Clavariaeforme DC. Эцидиальное спороношение на листьях боярышника (Crataegus), реже на груше. Телейстостадия на обыкновенном можжевельнике.
Triphragmium ulmariae Link.— возбудитель ржавчины лабазника вязолистного (Filipendula ulmaria).
Однохозяйный паразит с неполным циклом развития (отсутствуют эцидии). Спермогонии образуются вместе с первичными уредоложами.
Телейтоспоры бородавчатые на ножках, трехклеточные, с расположением клеток по углам треугольника: вверху две клетки рядом, внизу одна. Близкий вид Тг. filipendula Passer, имеет гладкие телейтоспоры, вызывает ржавчину лабазника шестилепестного (Filipendula hexapetala).
Phragmidium rubi — idael Karst. Телейтоспоры многоклеточные, сложены из 7—8 клеток, приплюснутые в вертикальном направлении, на длинной ножке, с утолщенной оболочкой. Эцидиальное спороношение типа цеома, без перидия, но с парафизами. Однохозяйный паразит, возбудитель ржавчины малины. Phr. disciflorum James.— возбудитель ржавчины роз; Phr. potentillae Karst.— возбудитель ржавчины лапчатки (Potentillae agrentea). Телейтоспоры преимущественно 5—6-клеточные.
Порядок Ржавчинные (Uredinales)
Грибы, относящиеся к этому порядку, вызывают заболевания многих растений. Симптомы поражения растений — пятна или полосы обычно ржаво-бурого цвета (табл. 54).
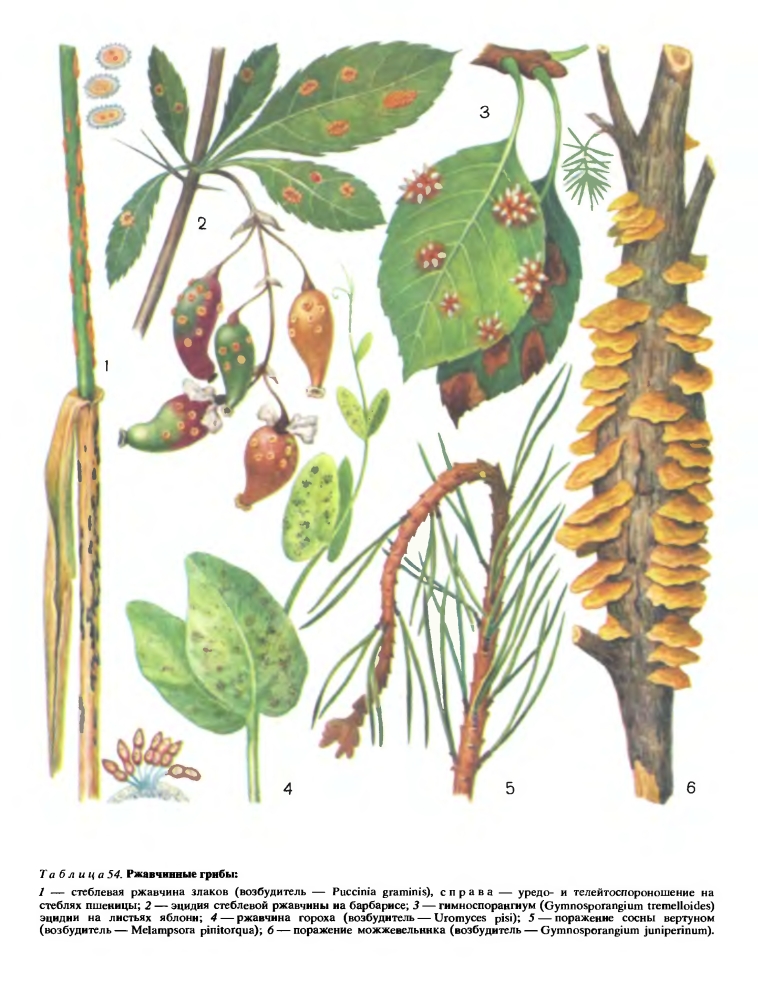
Отсюда название всей группы грибов этого порядка — ржавчинные грибы. Ржавчина хлебных злаков и других сельскохозяйственных растений известна с древних времен. Ряд писателей, живших до нашей эры, указывают на широкое распространение ржавчины хлебов в Египте, Греции, Риме и других древних государствах. Есть упоминания и о появлении ржавчины во времена удельной Руси. Опустошительные эпифитотии (массовые очаги) этой болезни на хлебах с древних времен до наших дней привлекли внимание людей к выяснению причин гибели растений при поражении ржавчиной. И хотя древние христиане ежегодно совершали молитвенные обряды в честь бога Робигуса, это не мешало им делать наблюдения в природе о характере заболевания на хлебах. Древние римляне задумывались над тем, может ли ржавчипа возникать в результате охлаждения растений или излишнего прогревания солнечными лучами осевших на листьях капель росы. Было замечено также, что растения в низинах поражаются ржавчиной сильнее и что разные сорта растений страдают от этой болезни в разной степени.
Внимание людей к ржавчине было обусловлено теми опустошительными потерями, которые наблюдались при массовом появлении ее на хлебах.
Ржавчинные грибы паразитируют на высших сосудистых растениях из многих семейств. Отличительной чертой этой группы грибов является наличие в их цикле развития нескольких различных по форме и функциям спороношений. Эти спороношения называются стадия ми развития гриба. Разные виды спор могут развиваться как на одном растении (однохоэяинные виды), так и на разных растениях (разнохозяинные виды).
При развитии ржавчины на растении обычно происходит местное, локальное заражение, т. е. поражение паблюдается па небольшом участке, куда попала одна спора гриба. У небольшого числа видов мицелий может пронизывать все растение и давать так называемое диффузное поражение всего растения. Для ржавчинных грибов характерно также массовое рассеивание спор. Оседая на растениях, споры ржавчины, переносимые ветром на многие километры, вызывают множество местных очагов инфекции на растении одного и того же вида.
Прорастающая спора ржавчинного гриба дает так называемую ростковую трубку, которая чаще всего через устьица проникает в ткани растения-хозяина. Мицелий гриба растет и распространяется внутри тканей растения, посылая в клетки гаустории, с помощью которых происходит питание гриба. Гифы ржавчинных грибов распространяются по межклетникам растения-хозяина. Контакт между цитоплазмой хозяина и паразита осуществляется в кончиках гаусторий. Недавно было показано, что на конце гаустории, примыкающей к цитоплазме растения-хозяина, оболочки нет. В этом месте есть только цитоплазменная мембрана, через которую питательные вещества хозяина переходят в мицелий паразита.
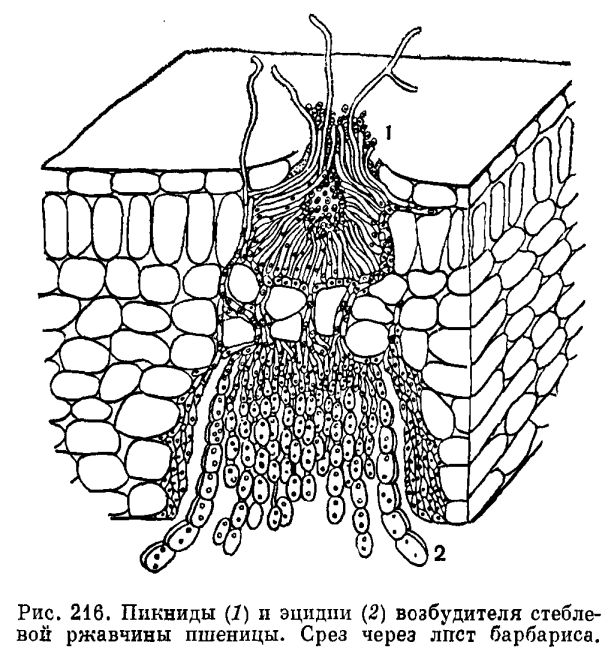
Для знакомства со сложным жизненным циклом ржавчинных грибов удобнее всего рассмотреть развитие возбудителя стеблевой (линейной) ржавчины злаков (Puccinia graminis). Этот гриб паразитирует на многих культурных и дикорастущих злаках, при массовом поражении он может полностью погубить урожай. Однако начало его развития отмечается не на злаках, а на растениях совсем другой группы, в данном случае на барбарисе. На листьях барбариса с верхней стороны в конце апреля — начале мая появляются оранжевые пятна. Это спороношение гриба, возникшее на мицелии паразита, находящемся в растении. Спороношение вначале представляет собой клубок гиф, которые затем преобразуются в бутылковидные структуры, погруженные в ткани листа. На вершине этих образований есть отверстие, дно и края их выстланы мицелием гриба, а в полости образуются специальные удлиненные клетки, отделяющие мелкие одноклеточные споры. Эти споры называют спермациями или пикноспорами, а бутылковидные структуры — спермогониями или пикнидами (рис. 216). Спермации и спермогонии возникают на одпоядерном мицелии гриба. При их созревании из отверстий выделяется сладковатая жидкость, привлекающая насекомых, переносящих споры гриба на другие листья барбариса.
Спермации и образующий их мицелий относятся к различным половым группам. Для дальнейшего развития гриба необходимо, чтобы содержимое двух различных спермациев соединилось друг с другом. Этот процесс по своей функции равнозначен половому процессу, свойственному остальным группам грибов. Соединение мицелиев различных полов может осуществляться насекомыми, которые приносят споры одного пола в спермогонии другого пола. Кроме того, на одном и том же листе прорастающие пикноспоры разных знаков могут подрастать друг к другу. Иногда спора одного полового знака может дать отросток в спермогоний, относящийся к другому полу. Во всех случаях происходит соединение ядер двух различных полов, в результате которого образуется двухядерный мицелий. Однако слияния ядер в этом случае не происходит, а начинается двухъядерная, или дикариофитная, стадия гриба.
Двухъядерный мицелий располагается под спермогониями внутри листа барбариса и в конце концов формирует на нижней поверхности его под спермогонием вместилище для двухъядерных спор — так называемый эцидий, внутри которого образуются эцидиоспоры (рис. 216). Эцидии имеют вид округлых или продолговатых чашечек, окруженных оболочкой, которая называется перидием и состоит из продолговатых бесцветных толстостенных клеток. На дне эцидия образуется слой цилиндрических клеток, от которого в виде цепочек отделяются эцидиоспоры. Между ними обычно находятся особые промежуточные клетки, которые впоследствии разрушаются и таким образом способствуют отделению эцидиоспор. Эцидиоспоры обычно округлой формы, одноклеточные и окрашены в ярко-желтый цвет. После созревания эцидий вскрывается и эцидиоспоры высыпаются во внешнюю среду. Эцидиоспоры, как и эцидии, образуются двухъядерным мицелием.
Для дальнейшего развития гриба необходимо, чтобы эцидиоспоры попали на растения из семейства злаков. На них и начинается основное развитие паразита. После заражения на злаках образуется местный, растущий на небольшом участке ткани растения двухъядерный мицелий, на котором развиваются летние споры гриба, называемые уредоспорами. Они возникают в массе под эпидермисом листа или стебля, а затем в случае стеблевой ржавчины прорывают его, образуя порошащие продолговатые щели, наполненные спорами гриба. Уредоспоры обычно одноклеточные, яйцевидной формы, оранжевые, расположенные на бесцветных ножках (табл. 54). Характерной чертой этого вида спороношения является то, что в течение лета оно может дать несколько поколений уредоспор, которые, попадая на соседние растения при благоприятных внешних условиях, вызывают массовое поражение хлебов (эпифитотию). Споры гриба распространяются от растения к растению ветром. Они могут подниматься воздушными течениями на высоту 2000 м и более, переноситься ветром на тысячи километров и, попадая на восприимчивые растения, вызывать их заражение.
Распространение уредоспор на большие расстояния свойственно всем ржавчинным грибам. Их появление в воздухе учитывают с помощью специальных стекол-ловушек, смазанных вазелином или другим жиром. Появление спор ржавчины учитывается специальной службой во всех странах мира и помогает предсказать возможность появления болезни на хлебных злаках в определенных районах.
Интенсивное поражение растения ржавчиной приводит к недоразвитию его, задержке образования колосьев и, таким образом, к гибели урожая.
К концу лета на месте уредоспор на том же двухъядерном мицелии образуются двухклеточные с темной толстой оболочкой телейтоспоры гриба (табл. 54). У возбудителя стеблевой ржавчины они расположены на ножках в телейтоспороношениях (пустулах), прорывающих эпидермис. Телейтоспоры служат для перезимовки гриба. Вначале они, так же как и образующий их мицелий, содержат два неслившихся ядра. Они зимуют на стерне злаков и способны оставаться живыми в течение всей зимы под снегом. Весной в спорах происходит слияние двух ядер (кариогамия), т. е. идет настоящий половой процесс. В результате этого слияния образуется ядро с двойным набором хромосом — диплоидное ядро. Этот процесс происходит до прорастания спор. Затем диплоидное ядро делится дважды, в результате чего образуются четыре гаплоидных (с единичным набором хромосом) ядра. На каждой клетке телейтоспоры развивается базидия — бесцветная клетка, разделенная перегородками на 4 части. От каждой части отходит бесцветный, утоньшающийся к концу вырост — стеригма, на кончике которой образуется базидиоспора, содержащая одно гаплоидное ядро. Для продолжения развития гриба необходимо, чтобы базидиоспора попала на растение барбариса. При прорастании на листе барбариса из каждой базидиоспоры развивается спермогоний, или пикнида. Поскольку базидиоспоры относятся к двум половым группам (две к одному полу, а две к другому), спермогонии также относятся к двум половым группам.
Таким образом, полный жизненный цикл возбудителя стеблевой ржавчины складывается из пяти следующих друг за другом спороношений:
Виды ржавчинных грибов, имеющих в своем развитии все указанные типы спороношений, относят к формам с полным циклом развития. У многих ржавчинных грибов некоторые типы спороношений отсутствуют. Их называют неполными формами. Как указывалось, эти спороношения могут развиваться на одном растении у однохозяинных видов и на различных растениях у разнохозяинных видов. Следовательно, вид Puccinia graminis относится к полным разнохозяинным видам ржавчинных грибов.
Важной особенностью ржавчинных грибов является приуроченность их к паразитированию на определенных растениях-хозяевах. Например, Puccinia graminis состоит из отдельных специализированных форм, которые различаются в основном по паразитированию на отдельных родах семейства злаков. Известны специализированные формы, приуроченные к пшенице (стеблевая ржавчина пшеницы), ржи (стеблевая ржавчина ржи), овсу (стеблевая ржавчина овса). Некоторые специализированные формы поражают дикорастущие злаки — полевицу, мятлик, тимофеевку, вейник и др.
Специализированные формы ржавчинных грибов, в свою очередь, делят на физиологические расы, отличающиеся друг от друга способностью поражать определенные сорта одного и того же вида растения-хозяина. Так, у возбудителя стеблевой ржавчины пшеницы к настоящему времени выявлено свыше 300 физиологических рас. Во всех районах мира ведется постоянный учет наличия тех или иных рас на посевах пшеницы и других культур, поражаемых ржавчиной. Эта работа нужна для того, чтобы своевременно выводить сорта, устойчивые к определенным расам возбудителей. Ржавчинные грибы отличаются значительной изменчивостью. В природе в результате полового процесса или мутаций происходят постоянные изменения в расовом составе паразитов в отдельных районах. Поэтому учет рас, поражающих сорта, ранее устойчивые к ним, проводится во всех странах мира. В нашей стране широкое распространение получили сорта пшеницы, выведенные академиком П. П. Лукьяненко, — Аврора, Кавказ, Безостая 1, отличающиеся относительной устойчивостью ко многим расам ржавчинных грибов.
Выведение устойчивых сортов — основной способ борьбы с болезнями, причиняемыми ржавчинными грибами. К другим защитным мероприятиям относятся уничтожение промежуточных хозяев гриба (например, барбариса в случае со стеблевой ржавчиной) и недопущение посевов этих растений рядом с посевами зерновых культур.
Агротехнические мероприятия, ограничивающие развитие заболеваний растений ржавчиной, состоят в улучшенном содержании посевов зерновых культур. Химический способ борьбы (обработка посевов ядохимикатами) экономически выгоден только для уничтожения ржавчины на многолетних культурах или для обработки ценного селекционного материала.
Распространение ржавчинных грибов связано с распространением растений, на которых они развиваются. Тесная связь ржавчины и растения обусловлена их совместной эволюцией. П. М. Жуковский, развивая взгляды Н. И. Вавилова, создал теорию сопряженной эволюции паразита и растения-хозяина. По этой теории, в центрах происхождения растений наряду с разнообразием форм растений наблюдается разнообразие форм паразита. Распространение растений по земному шару приводит к распространению рас паразита.
Порядок Uredinaies объединяет свыше 4000 видов. Их систематика основывается на строении телейтоспор, поскольку именно в них происходит половой процесс — слияние ядер, образование диплоидного ядра и последующее редукционное деление. Кроме того, телейтоспоры известны как у полных, так и у неполных видов ржавчинных грибов. Телейтоспоры отличаются большим разнообразием. Они могут располагаться на ножке или быть лишенными ее, могут развиваться отдельно друг от друга или в слизистых скоплениях, могут соединяться боковыми сторонами, объединяться в колонки и т. д. У грибов различных видов они отличаются по цвету (от почти бесцветных до темно-коричневых), размерам, форме, поверхности оболочки и т. п. Во всех случаях телейтоспоры служат зимующей стадией гриба.
Порядок ржавчинных грибов делят на два семейства — мелампсоровых (Melampsoraceae) и пукциниевых (Pucciniaceae).
Жизнь растений: в 6-ти томах. — М.: Просвещение. Под редакцией А. Л. Тахтаджяна, главный редактор чл.-кор. АН СССР, проф. А.А. Федоров . 1974 .
Размножение и особенности жизненного цикла ржавчинных грибов
Ржавчинные грибы — это высшие многоклеточные грибы отдела Базидиомицеты, облигатные паразиты (живут в организме хозяина в анаэробных условиях) покрытосеменных, голосеменных и папоротникообразных растений. Ржавчинные вызывают заболевания множества растений. Признаки заражения — это появление пятен или линий ржавого цвета. Так и сформировалось название порядка — ржавчинные грибы.
Характеристика и особенности жизненного цикла ржавчинных грибов

Порядок ржавчинные включает примерно 4000 видов грибов, которые имеют специальные органы размножения — базидии, они состоят из одноклеточных спор. Спора, попадая во внутреннюю среду растения, дает новый росток, который при благоприятных условиях созревает до мицелия.
Спора созревает на протяжении пяти последовательных стадий:
- пикноконидии и эцидиоспоры – развиваются в весенний период;
- уредоспоры — летние споры;
- телейтоспоры и базидиоспоры — осенне-зимний период жизненного цикла.
На них возложены определенные задачи: поражение растений происходит при помощи эцидиоспор, уредоспор или базидиоспор, пережидание зимнего периода возможно в виде телейтоспор. На этапе телейтоспоры грибы для растений не опасны, они просто дальше преобразуются в базидиоспоры. Ржавчинные грибы иногда пережидают холода внутри растений, живущих более года, а по весне процесс снова начинается сначала.
Грибы, которые в своем развитии прошли все стадии, являются грибами с полным циклом развития. У представителей с неполным циклом развития отсутствуют отдельные стадии. Эти виды ржавчинных можно встретить в зоне умеренного климата, поскольку у них непродолжительный вегетационный период.
Род пукциния
Род пукциния самый многочисленный в порядке ржавчинных грибов: насчитывается примерно 1800 видов. Есть представители, живущие с одним хозяином или разными, с полным циклом развития или неполным. Паразитируют на злаковых, многолетних травах, поражают однодольные, астровые растения. Например, гриб рода пукциниевых с разными хозяевами — Puccinia graminis (вызывает стеблевую ржавчину злаковых). В жизненном цикле гриба принимают участие: представитель из семейства Барбарисовые и злаковые растения.
Спороносная стадия развития у ржавчинных грибов располагается на листьях и стеблях растений. На зараженных частях возникают ржаво-бурые пятна, которые расположены в виде линий. За лето они дозревают, высыхают и на них образуются трещины, через которые высыпаются споры, легко распространяющиеся на рядом расположенные растения.

Представитель рода пукциния
Размножение ржавчинных грибов
Каждая из спор является двухъядерной – это уредоспора, внутри оранжевого цвета, есть оболочка с шипами и порами. Они имеют овальную форму и располагаются на ножнах, являясь окончаниями нитевидных образований грибов. Уредоспоры прорастают очень быстро, попадая на другое растение, проникают внутрь, разрастаются там, формируя мицелий.
Споры выделяются за летний период несколько раз, поэтому ржавчинные грибы так легко поражают большую площадь растений. Питательные вещества, необходимые для роста ржавчины, поступают из клеток хозяина, что постепенно истощает его организм.
На месте ржаво-желтых пятен в начале осени появляются темные. В них содержатся споры, которые прорастут лишь весной. Это зимние споры – телейтоспоры, они двухклеточные с прочной оболочкой, защищающей споры в зимний период.
Все телейтоспоры пукциниевых грибов вначале имеют по два ядра. На стадии созревания ядра объединяются в одно – диплоидное ядро. С приходом весны на останках злаковых растений спора начинает прорастать и выбрасывает две нити, куда перемещаются ядра. Ядра, с двойным набором хромосом, начинают делятся с помощью непрямого деления, так образуется по четыре клетки с каждой нити, уже с гаплоидными набором хромосом (базидиальные клетки). Базидиоспоры пукциниевых грибов разлетаются по ветру и могут прорастать лишь при попадании на листья барбариса.
На верхней поверхности листьев барбариса появляются темные точки, это пикнидии. Они отделяют от себя споры – спермации (с гаплоидными ядрами), которые нужны для перехода в двухъядерную стадию.
На нижней стороне листа барбариса образуются – эцидии, в которых находятся двухъядерные эцидиоспоры. Эцидиоспоры с помощью ветра заносятся на стебли и листья злаковых, где вскоре прорастают. Так новообразованный гриб снова производит летние споры – уредоспоры и процесс начинается сначала.
Спороносная стадия развития у ржавчинных грибов приводит к массовому распространению грибов, и они могут уничтожать целые поля злаковых растений. Поражение растений ржавчиной наносит большой урон урожаю, для борьбы с грибами разработаны устойчивые сорта злаков, проводят обработку зерновых фунгицидами, лущение стерни. При поражении деревьев, кустарников оптимально, прежде всего, сжигать поражённые ветки и листья.
Источник https://www.activestudy.info/poryadok-rzhavchinnye-uredinales/
Источник https://dic.academic.ru/dic.nsf/enc_biology/1666/%D0%9F%D0%BE%D1%80%D1%8F%D0%B4%D0%BE%D0%BA
Источник https://animals-world.ru/rzhavchinnye-griby/


